Direct Approach to FDR and q-values
Overview
Teaching: 20 min
Exercises: 40 minQuestions
How can you control false discovery rates when you don’t have an a priori error rate?
Objectives
Apply the Storey correction to identify true negatives.
Explain the difference between the Storey correction and Benjamini-Hochberg approach.
Direct Approach to FDR and q-values (Advanced)
Here we review the results described by John D. Storey in J. R. Statist. Soc. B (2002). One major distinction between Storey’s approach and Benjamini and Hochberg’s is that we are no longer going to set a $\alpha$ level a priori. Because in many high-throughput experiments we are interested in obtaining some list for validation, we can instead decide beforehand that we will consider all tests with p-values smaller than 0.01. We then want to attach an estimate of an error rate. Using this approach, we are guaranteed to have $R>0$. Note that in the FDR definition above we assigned $Q=0$ in the case that $R=V=0$. We were therefore computing:
\[\mbox{FDR} = E\left( \frac{V}{R} \mid R>0\right) \mbox{Pr}(R>0)\]In the approach proposed by Storey, we condition on having a non-empty list, which implies $R>0$, and we instead compute the positive FDR
\[\mbox{pFDR} = E\left( \frac{V}{R} \mid R>0\right)\]A second distinction is that while Benjamini and Hochberg’s procedure controls under the worst case scenario, in which all null hypotheses are true ( $m=m_0$ ), Storey proposes that we actually try to estimate $m_0$ from the data. Because in high-throughput experiments we have so much data, this is certainly possible. The general idea is to pick a relatively high value p-value cut-off, call it $\lambda$, and assume that tests obtaining p-values > $\lambda$ are mostly from cases in which the null hypothesis holds. We can then estimate $\pi_0 = m_0/m$ as:
\[\hat{\pi}_0 = \frac{\#\left\{p_i > \lambda \right\} }{ (1-\lambda) m }\]There are more sophisticated procedures than this, but they follow the same general idea. Here is an example setting $\lambda=0.1$. Using the p-values computed above we have:
library(genefilter) ##rowttests is here
set.seed(1)
alpha <- 0.05
N <- 12
##Define groups to be used with rowttests
g <- factor( c(rep(0, N), rep(1, N)) )
# re-create p-values from earlier if needed
population <- unlist(read.csv(file = "../data/femaleControlsPopulation.csv"))
m <- 10000
p0 <- 0.90
m0 <- m*p0
m1 <- m-m0
nullHypothesis <- c( rep(TRUE,m0), rep(FALSE,m1))
delta <- 3
B <- 1000 ##number of simulations. We should increase for more precision
res <- replicate(B, {
controls <- matrix(sample(population, N*m, replace=TRUE),nrow=m)
treatments <- matrix(sample(population, N*m, replace=TRUE),nrow=m)
treatments[which(!nullHypothesis),]<-treatments[which(!nullHypothesis),]+delta
dat <- cbind(controls,treatments)
pvals <- rowttests(dat,g)$p.value
##then the FDR
calls <- p.adjust(pvals,method="fdr") < alpha
R=sum(calls)
Q=ifelse(R>0,sum(nullHypothesis & calls)/R,0)
return(c(R,Q))
}
)
Qs <- res[2,]
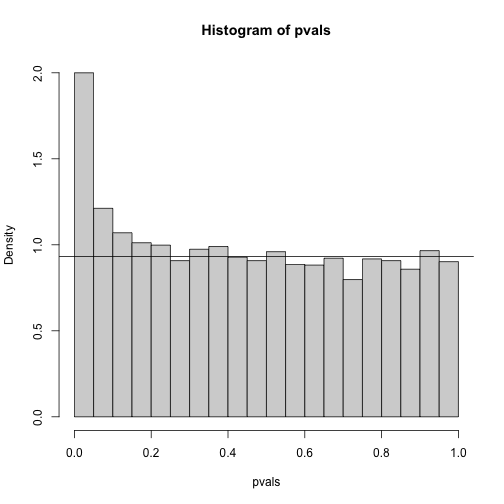
hist(pvals,breaks=seq(0,1,0.05),freq=FALSE)
## Error in hist(pvals, breaks = seq(0, 1, 0.05), freq = FALSE): object 'pvals' not found
lambda = 0.1
pi0=sum(pvals> lambda) /((1-lambda)*m)
## Error in eval(expr, envir, enclos): object 'pvals' not found
abline(h= pi0)
## Error in int_abline(a = a, b = b, h = h, v = v, untf = untf, ...): object 'pi0' not found
print(pi0) ##this is close to the trye pi0=0.9
## Error in print(pi0): object 'pi0' not found
With this estimate in place we can, for example, alter the Benjamini and Hochberg procedures to select the $k$ to be the largest value so that:
\[\hat{\pi}_0 p_{(i)} \leq \frac{i}{m}\alpha\]However, instead of doing this, we compute a q-value for each test. If a feature resulted in a p-value of $p$, the q-value is the estimated pFDR for a list of all the features with a p-value at least as small as $p$.
In R, this can be computed with the qvalue function in the qvalue package:
library(qvalue)
res <- qvalue(pvals)
## Error in qvalue(pvals): object 'pvals' not found
qvals <- res$qvalues
## Error in res$qvalues: $ operator is invalid for atomic vectors
plot(pvals, qvals)
## Error in h(simpleError(msg, call)): error in evaluating the argument 'x' in selecting a method for function 'plot': object 'pvals' not found
we also obtain the estimate of $\hat{\pi}_0$:
res$pi0
## Error in res$pi0: $ operator is invalid for atomic vectors
This function uses a more sophisticated approach at estimating $\pi_0$ than what is described above.
Note on estimating $\pi_0$
In our experience the estimation of $\pi_0$ can be unstable and adds a step of uncertainty to the data analysis pipeline. Although more conservative, the Benjamini-Hochberg procedure is computationally more stable.
Exercises
In the following exercises, we will define error controlling procedures for experimental data. We will make a list of genes based on q-values. We will also assess your understanding of false positives rates and false negative rates by asking you to create a Monte Carlo simulation.
Exercise 1
Load the gene expression data:
library(GSE5859Subset)
data(GSE5859Subset)
We are interested in comparing gene expression between the two groups defined in the sampleInfo table. Compute a p-value for each gene using the functionrowttestsfrom the genefilter package.
library(genefilter)
?rowttestsHow many genes have p-values smaller than 0.05?
Solution
g <- sampleInfo$group
pvals <- rowttests(geneExpression, factor(g))$p.value
sum(pvals < 0.05)
Exercise 2
Apply the Bonferroni correction to achieve a FWER of 0.05. How many genes are called significant under this procedure?
Solution
m <- 8793
sum(pvals < (0.05/m))
Exercise 3
The FDR is a property of a list of features, not each specific feature. The q-value relates FDR to individual features. To define the q-value, we order features we tested by p-value, then compute the FDRs for a list with the most significant, the two most significant, the three most significant, etc. The FDR of the list with the, say, m most significant tests is defined as the q-value of the m-th most significant feature. In other words, the q-value of a feature, is the FDR of the biggest list that includes that gene. In R, we can compute q-values using the p.adjust function with the FDR option. Read the help file for p.adjust and compute how many genes achieve a q-value < 0.05 for our gene expression dataset.
Solution
pvals_adjust <- p.adjust(pvals, method = 'fdr')
sum(pvals_adjust < 0.05)
Exercise 4
Now use the qvalue function, in the Bioconductor qvalue package, to estimate q-values using the procedure described by Storey. How many genes have q-values below 0.05?
Solution
res <- qvalue(pvals)
sum(res$qvalues < 0.05)
Exercise 5
Read the help file for qvalue and report the estimated proportion of genes for which the null hypothesis is true π0 = m0/m
Solution
res$pi0
Exercise 6
The number of genes passing the q-value < 0.05 threshold is larger with the q-value function than the p.adjust difference. Why is this the case? Make a plot of the ratio of these two estimates to help answer the question.
A) One of the two procedures is flawed.
B) The two functions are estimating different things.
C) The qvalue function estimates the proportion of genes for which the null hypothesis is true and provides a less conservative estimate.
D) The qvalue function estimates the proportion of genes for which the null hypothesis is true and provides a more conservative estimate.Solution
plot(pvals_adjust, res$qvalues, xlab = 'fdr', ylab = 'qval')
abline(0,1)
The qvalue function estimates the proportion of genes for which the null hypothesis is true and provides a less conservative estimate (choice C).
Exercise 7
This exercise and the remaining ones are more advanced. Create a Monte Carlo Simulation in which you simulate measurements from 8,793 genes for 24 samples, 12 cases and 12 controls. Then for 100 genes create a difference of 1 between cases and controls. You can use the code provided below. Run this experiment 1,000 times with a Monte Carlo simulation. For each instance, compute p-values using a t-test and keep track of the number of false positives and false negatives. Compute the false positive rate and false negative rates if we use Bonferroni, q-values from p.adjust, and q-values from qvalue function. Set the seed to 1 for all three simulations. What is the false positive rate for Bonferroni? What are the false negative rates for Bonferroni?
n <- 24 m <- 8793 mat <- matrix(rnorm(n*m),m,n) delta <- 1 positives <- 500 mat[1:positives,1:(n/2)] <- mat[1:positives,1:(n/2)]+deltaSolution
g <- c(rep(0,12),rep(1,12)) m <- 8793 B <- 1000 m1 <- 500 N <- 12 m0 <- m-m1 nullHypothesis <- c(rep(TRUE,m0),rep(FALSE,m1)) delta <- 1 set.seed(1) res <- replicate(B, { controls <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment[!nullHypothesis,] <- treatment[!nullHypothesis,] + delta dat <- cbind(controls, treatment) pvals <- rowttests(dat, factor(g))$p.value calls <- pvals < (0.05/m) R <- sum(calls) V <- sum(nullHypothesis & calls) fp <- sum(nullHypothesis & calls)/m0 # false positive fn <- sum(!nullHypothesis & !calls)/m1 # false negative return(c(fp,fn)) }) res<-t(res) head(res) mean(res[,1]) # false positive rate mean(res[,2]) # false negative rate
Exercise 8
What are the false positive rates for p.adjust?
What are the false negative rates for p.adjust?Solution
g <- c(rep(0,12),rep(1,12)) m <- 8793 B <- 1000 m1 <- 500 N <- 12 m0 <- m-m1 nullHypothesis <- c(rep(TRUE,m0),rep(FALSE,m1)) delta <- 1 set.seed(1) res <- replicate(B, { controls <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment[!nullHypothesis,] <- treatment[!nullHypothesis,] + delta dat <- cbind(controls, treatment) pvals <- rowttests(dat, factor(g))$p.value pvals_adjust <- p.adjust(pvals, method = 'fdr') calls <- pvals_adjust < 0.05 R <- sum(calls) V <- sum(nullHypothesis & calls) fp <- sum(nullHypothesis & calls)/m0 # false positive fn <- sum(!nullHypothesis & !calls)/m1 # false negative return(c(fp,fn)) }) res <- t(res) head(res) mean(res[,1]) # false positive rate mean(res[,2]) # false negative rate
Exercise 9
What are the false positive rates for qvalues?
What are the false negative rates for qvalues?Solution
g <- c(rep(0,12),rep(1,12)) m <- 8793 B <- 1000 m1 <- 500 N <- 12 m0 <- m-m1 nullHypothesis <- c(rep(TRUE,m0),rep(FALSE,m1)) delta <- 1 set.seed(1) res <- replicate(B, { controls <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment <- matrix(rnorm(N*m),nrow = m, ncol = N) treatment[!nullHypothesis,] <- treatment[!nullHypothesis,] + delta dat <- cbind(controls, treatment) pvals <- rowttests(dat, factor(g))$p.value qvals <- qvalue(pvals)$qvalue calls <- qvals < 0.05 R <- sum(calls) V <- sum(nullHypothesis & calls) fp <- sum(nullHypothesis & calls)/m0 # false positive fn <- sum(!nullHypothesis & !calls)/m1 # false negative return(c(fp,fn)) }) res <- t(res) head(res) mean(res[,1]) # false positive rate mean(res[,2]) # false negative rate
Key Points
The Storey correction makes different assumptions that Benjamini-Hochberg. It does not set a priori alpha levels, but instead estimates the number of true null hypotheses from a given data set.
The Storey correction is less computationally stable than Benjamini-Hochberg.